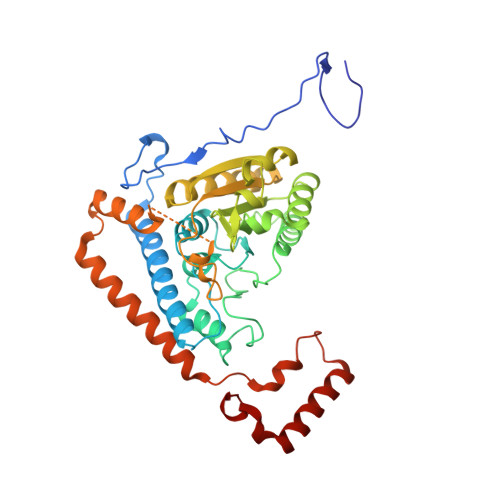
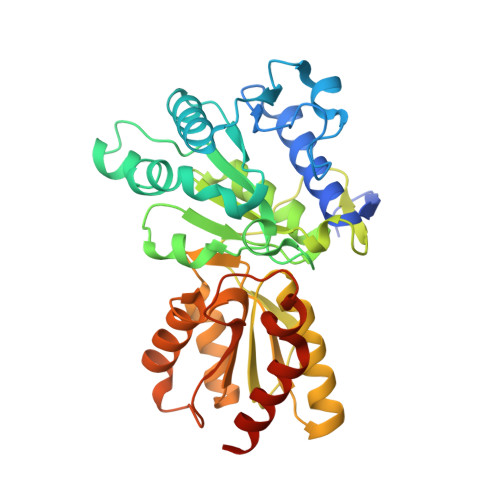
The Two Active Sites in Human Branched-Chain Alpha- Keto Acid Dehydrogenase Operate Independently without an Obligatory Alternating-Site Mechanism.
Li, J., Machius, M., Chuang, J.L., Wynn, R.M., Chuang, D.T.(2007) J Biological Chem 282: 11904
- PubMed: 17329260 Search on PubMed
- DOI: https://doi.org/10.1074/jbc.M610843200
- Primary Citation Related Structures:
2J9F - PubMed Abstract:
A long standing controversy is whether an alternating activesite mechanism occurs during catalysis in thiamine diphosphate (ThDP)-dependent enzymes. We address this question by investigating the ThDP-dependent decarboxylase/dehydrogenase (E1b) component of the mitochondrial branched-chain alpha-keto acid dehydrogenase complex (BCKDC). Our crystal structure reveals that conformations of the two active sites in the human E1b heterotetramer harboring the reaction intermediate are identical. Acidic residues in the core of the E1b heterotetramer, which align with the proton-wire residues proposed to participate in active-site communication in the related pyruvate dehydrogenase from Bacillus stearothermophilus, are mutated. Enzyme kinetic data show that, except in a few cases because of protein misfolding, these alterations are largely without effect on overall activity of BCKDC, ruling out the requirement of a proton-relay mechanism in E1b. BCKDC overall activity is nullified at 50% phosphorylation of E1b, but it is restored to nearly half of the pre-phosphorylation level after dissociation and reconstitution of BCKDC with the same phosphorylated E1b. The results suggest that the abolition of overall activity likely results from the specific geometry of the half-phosphorylated E1b in the BCKDC assembly and not due to a disruption of the alternating active-site mechanism. Finally, we show that a mutant E1b containing only one functional active site exhibits half of the wild-type BCKDC activity, which directly argues against the obligatory communication between active sites. The above results provide evidence that the two active sites in the E1b heterotetramer operate independently during the ThDP-dependent decarboxylation reaction.
- Department of Biochemistry, University of Texas Southwestern Medical Center, Dallas, Texas 75390, USA.
Organizational Affiliation: