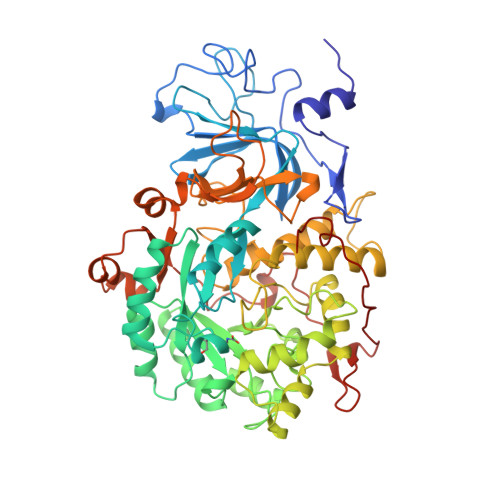
Structures of Cys319 variants and acetohydroxamate-inhibited Klebsiella aerogenes urease.
Pearson, M.A., Michel, L.O., Hausinger, R.P., Karplus, P.A.(1997) Biochemistry 36: 8164-8172
- PubMed: 9201965 Search on PubMed
- DOI: https://doi.org/10.1021/bi970514j
- Primary Citation Related Structures:
1FWA, 1FWB, 1FWC, 1FWD, 1FWE, 1FWF, 1FWG, 1FWH, 1FWJ - PubMed Abstract:
Cys319 is located on a mobile flap covering the active site of Klebsiella aerogenes urease but does not play an essential role in catalysis. Four urease variants altered at position C319 range from having high activity (C319A) to no measurable activity (C319Y), indicating Cys is not required at this position, but its presence is highly influential [Martin, P. R., & Hausinger, R. P. (1992) J. Biol. Chem. 267, 20024-20027]. Here, we present 2.0 A resolution crystal structures of C319A, C319S, C319D, and C319Y proteins and the C319A variant inhibited by acetohydroxamic acid. These structures show changes in the hydration of the active site nickel ions and in the position and flexibility of the active site flap. The C319Y protein exhibits an alternate conformation of the flap, explaining its lack of activity. The changes in hydration and conformation suggest that there are suboptimal protein-solvent and protein-protein interactions in the empty urease active site which contribute to urease catalysis. Specifically, we hypothesize that the suboptimal interactions may provide a significant source of substrate binding energy, and such hidden energy may be a common phenomenon for enzymes that contain mobile active site loops and undergo an induced fit. The acetohydroxamic acid-bound structure reveals a chelate interaction similar to those seen in other metalloenzymes and in a small molecule nickel complex. The inhibitor binding mode supports the proposed mode of urea binding. We complement these structural studies with extended functional studies of C319A urease to show that it has enhanced stability and resistance to inhibition by buffers containing nickel ions. The near wild-type activity and enhanced stability of the C319A variant make it useful for further studies of urease structure-function relationships.
- Section of Biochemistry, Cornell University, Ithaca, New York 14853, USA.
Organizational Affiliation: