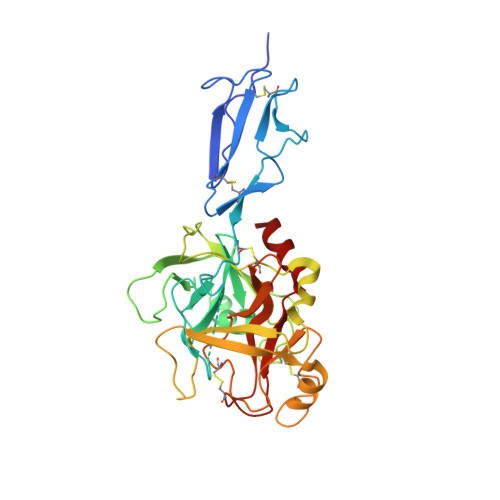
The structure of MBL-associated serine protease-2 reveals that identical substrate specificities of C1s and MASP-2 are realized through different sets of enzyme-substrate interactions
Harmat, V., Gal, P., Kardos, J., Szilagyi, K., Ambrus, G., Vegh, B., Naray-Szabo, G., Zavodsky, P.(2004) J Mol Biol 342: 1533-1546
- PubMed: 15364579 Search on PubMed
- DOI: https://doi.org/10.1016/j.jmb.2004.07.014
- Primary Citation Related Structures:
1Q3X - PubMed Abstract:
A family of serine proteases mediates the proteolytic cascades of several defense mechanisms in vertebrates, such as the complement system, blood coagulation and fibrinolysis. These proteases usually form large complexes with other glycoproteins. Their common features are their modular structures and restricted substrate specificities. The lectin pathway of complement, where mannose-binding lectin (MBL) recognizes the carbohydrate structures on pathogens, is activated by mannose-binding lectin-associated serine protease-2 (MASP-2). We present the 2.25A resolution structure of the catalytic fragment of MASP-2 encompassing the second complement control protein module (CCP2) and the serine protease (SP) domain. The CCP2 module stabilizes the structure of the SP domain as demonstrated by differential scanning calorimetry measurements. The asymmetric unit contains two molecules with different CCP-SP domain orientations, reflecting increased modular flexibility at the CCP2/SP joint. This flexibility may partly explain the ability of the MASP-2 dimer to perform all of its functions alone, whereas the same functions are mediated by the much larger C1r2-C1s2 tetramer in the C1 complex of the classical pathway. The main scaffold of the MASP-2 SP domain is chymotrypsin-like. Eight surface loops determine the S1 and other subsite specificities. Surprisingly, some surface loops of MASP-2, e.g. loop 1 and loop 2, which form the S1 pocket are similar to those of trypsin, and show significant differences if compared with those of C1s, indicating that the nearly identical substrate specificities of C1s and MASP-2 are realized through different sets of enzyme-substrate interactions.
- Protein Modeling Group, Hungarian Academy of Sciences, Eötvös Loránd University, Pázmány Péter sétány. 1A, H-1117 Budapest, Hungary.
Organizational Affiliation: