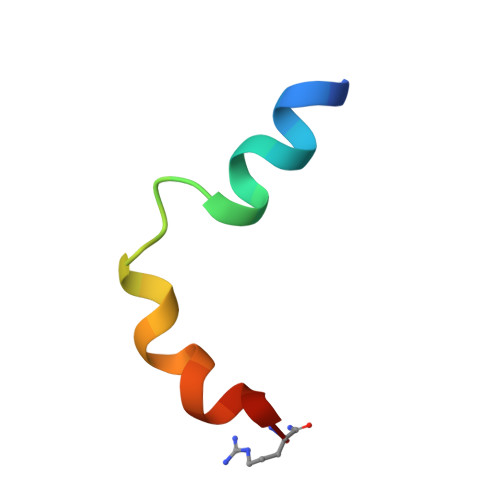
Determination of the structure of the N-terminal splice region of the cyclic AMP-specific phosphodiesterase RD1 (RNPDE4A1) by 1H NMR and identification of the membrane association domain using chimeric constructs.
Smith, K.J., Scotland, G., Beattie, J., Trayer, I.P., Houslay, M.D.(1996) J Biol Chem 271: 16703-16711
- PubMed: 8663181 Search on PubMed
- DOI: https://doi.org/10.1074/jbc.271.28.16703
- Primary Citation Related Structures:
1LOI - PubMed Abstract:
A 25-residue peptide representing the membrane targeting N-terminal splice region of the cyclic AMP phosphodiesterase RD1 (RNPDE4A1) was synthesized, and its structure was determined by 1H NMR. Two independently folding helical regions were identified, separated by a highly mobile "hinge" region. The first helical region was formed by an N-terminal amphipathic alpha-helix, and the second consisted of multiple overlapping turns and contained a distinct compact, hydrophobic, tryptophan-rich domain (residues 14-20). Chimeric molecules, formed between the N-terminal region of RD1 and the soluble bacterial protein chloramphenicol acetyltransferase, were used in an in vitro system to determine the features within the splice region that were required for membrane association. The ability of RD1-chloramphenicol acetyltransferase chimera to become membrane-associated was not affected by deletion of any of the following regions: the apolar section (residues 2-7) of the first helical region, the polar part of this region together with the hinge region (residues 8-13), or the polar end of the C-terminal helical region (residues 21-25). In marked contrast, deletion of the compact, hydrophobic tryptophan-rich domain (residues 14-20) found in the second helical region obliterated membrane association. Replacement of this domain with a hydrophobic cassette of seven alanine residues also abolished membrane association, indicating that membrane-association occurred by virtue of specific hydrophobic interactions with residues within the compact, tryptophan-rich domain. The structure of this domain is well defined in the peptide, and although the region is helical, both the backbone and the distribution of side chains are somewhat distorted as compared with an ideal alpha-helix. Hydrophobic interactions, such as the "stacked" rings of residues Pro14 and Trp15, stabilize this domain with the side chain of residue Leu16 adopting a central position, interacting with the side chains of all three tryptophan residues 15, 19, and 20. These bulky side chains thus form a hydrophobic cluster. In contrast, the side chain of residue Val17 is relatively exposed, pointing out from the opposite "face" of the peptide. Although it appears that this compact, tryptophan-rich domain is responsible for membrane association, at present the target site and hence the specific interactions involved in membrane targeting by the RD1 splice region remain unidentified.
- Molecular Pharmacology Group, Division of Biochemistry and Molecular Biology, Institute of Life and Biomedical Sciences, University of Glasgow, Glasgow G12 8QQ, Scotland.
Organizational Affiliation: