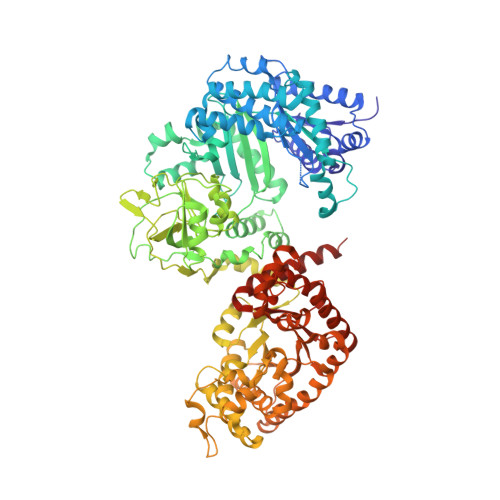
The 3.0 A Resolution Crystal Structure of Glycosomal Pyruvate Phosphate Dikinase from Trypanosoma Brucei
Cosenza, L.W., Bringaud, F., Baltz, T., Vellieux, F.M.D.(2001) J Mol Biology 318: 1417
- PubMed: 12083528 Search on PubMed
- DOI: https://doi.org/10.1016/s0022-2836(02)00113-4
- Primary Citation Related Structures:
2X0S - PubMed Abstract:
The crystal structure of the glycosomal enzyme pyruvate phosphate dikinase from the African protozoan parasite Trypanosoma brucei has been solved to 3.0 A resolution by molecular replacement. The search model was the 2.3 A resolution structure of the Clostridium symbiosum enzyme. Due to different relative orientations of the domains and sub-domains in the two structures, molecular replacement could be achieved only by positioning these elements (four bodies altogether) sequentially in the asymmetric unit of the P2(1)2(1)2 crystal, which contains one pyruvate phosphate dikinase (PPDK) subunit. The refined model, comprising 898 residues and 188 solvent molecules per subunit, has a crystallographic residual index Rf = 0.245 (cross-validation residual index Rfree = 0.291) and displays satisfactory stereochemistry. Eight regions, comprising a total of 69 amino acid residues at the surface of the molecule, are disordered in this crystal form. The PPDK subunits are arranged around the crystallographic 2-fold axis as a dimer, analogous to that observed in the C. symbiosum enzyme. Comparison of the two structures was carried out by superposition of the models. Although the fold of each domain or sub-domain is similar, the relative orientations of these constitutive elements are different in the two structures. The trypanosome enzyme is more "bent" than the bacterial enzyme, with bending increasing from the center of the molecule (close to the molecular 2-fold axis) towards the periphery where the N-terminal domain is located. As a consequence of this increased bending and of the differences in relative positions of subdomains, the nucleotide-binding cleft in the amino-terminal domain is wider in T. brucei PPDK: the N-terminal fragment of the amino-terminal domain is distant from the catalytic, phospho-transfer competent histidine 482 (ca 10 A away). Our observations suggest that the requirements of domain motion during enzyme catalysis might include widening of the nucleotide-binding cleft to allow access and departure of the AMP or ATP ligand.
- Laboratoire de Biophysique Moleculaire, Institut de Biologie Structurale J.-P. Ebel CEA CNRS UJF, Grenoble, France.
Organizational Affiliation: