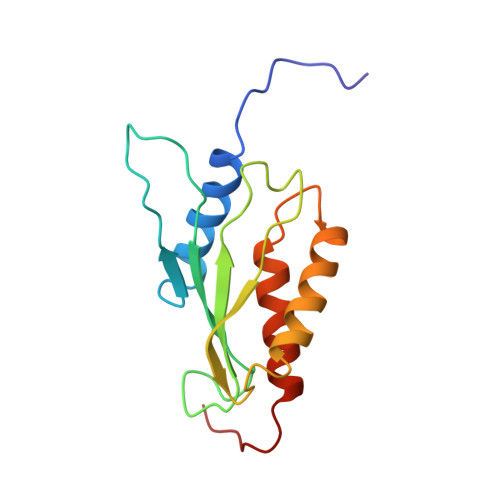
Solution structure of the RWD domain of the mouse GCN2 protein.
Nameki, N., Yoneyama, M., Koshiba, S., Tochio, N., Inoue, M., Seki, E., Matsuda, T., Tomo, Y., Harada, T., Saito, K., Kobayashi, N., Yabuki, T., Aoki, M., Nunokawa, E., Matsuda, N., Sakagami, N., Terada, T., Shirouzu, M., Yoshida, M., Hirota, H., Osanai, T., Tanaka, A., Arakawa, T., Carninci, P., Kawai, J., Hayashizaki, Y., Kinoshita, K., Guntert, P., Kigawa, T., Yokoyama, S.(2004) Protein Sci 13: 2089-2100
- PubMed: 15273307 Search on PubMedSearch on PubMed Central
- DOI: https://doi.org/10.1110/ps.04751804
- Primary Citation Related Structures:
1UKX - PubMed Abstract:
GCN2 is the alpha-subunit of the only translation initiation factor (eIF2alpha) kinase that appears in all eukaryotes. Its function requires an interaction with GCN1 via the domain at its N-terminus, which is termed the RWD domain after three major RWD-containing proteins: RING finger-containing proteins, WD-repeat-containing proteins, and yeast DEAD (DEXD)-like helicases. In this study, we determined the solution structure of the mouse GCN2 RWD domain using NMR spectroscopy. The structure forms an alpha + beta sandwich fold consisting of two layers: a four-stranded antiparallel beta-sheet, and three side-by-side alpha-helices, with an alphabetabetabetabetaalphaalpha topology. A characteristic YPXXXP motif, which always occurs in RWD domains, forms a stable loop including three consecutive beta-turns that overlap with each other by two residues (triple beta-turn). As putative binding sites with GCN1, a structure-based alignment allowed the identification of several surface residues in alpha-helix 3 that are characteristic of the GCN2 RWD domains. Despite the apparent absence of sequence similarity, the RWD structure significantly resembles that of ubiquitin-conjugating enzymes (E2s), with most of the structural differences in the region connecting beta-strand 4 and alpha-helix 3. The structural architecture, including the triple beta-turn, is fundamentally common among various RWD domains and E2s, but most of the surface residues on the structure vary. Thus, it appears that the RWD domain is a novel structural domain for protein-binding that plays specific roles in individual RWD-containing proteins.
- RIKEN Genomic Sciences Center, Tsurumi, Yokohama, Japan.
Organizational Affiliation: