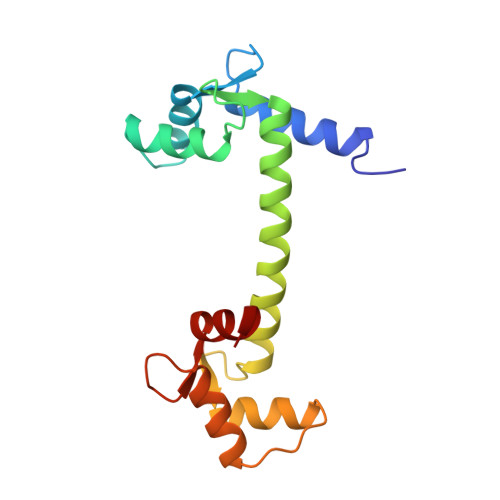
Structure of the recombinant Paramecium tetraurelia calmodulin at 1.68 A resolution.
Ban, C., Ramakrishnan, B., Ling, K.Y., Kung, C., Sundaralingam, M.(1994) Acta Crystallogr D Biol Crystallogr 50: 50-63
- PubMed: 15299476 Search on PubMed
- DOI: https://doi.org/10.1107/S0907444993007991
- Primary Citation Related Structures:
1OSA - PubMed Abstract:
The crystal structure of the recombinant calmodulin from Paramecium tetraurelia (rPCaM, M(r) = 16 700, 148 residues) has been determined at 1.68 A resolution. X-ray intensity data were collected at 263 K using a Siemens-Nicolet area detector and Cu Kalpha radiation from a rotating-anode source. A total of 35 936 observations were processed with XENGEN1.3 and scaled to yield 16 255 unique reflections with R(symm)(I) of 4.1%. The crystals are triclinic, with unit-cell dimensions a = 29.89, b = 53.42, c = 25.35 A, alpha = 93.67, beta = 96.88, gamma = 89.24 degrees, space group P1, with one molecule in the unit cell. The atomic coordinates of the wild-type Paramecium calmodulin (PCaM) studied in our laboratory provided the starting model. Refinement of the structure by X-PLOR and refitting it into omit maps yielded an R value of 0.194 for 15 965 reflections greater than 3sigma(F) in the 6.0-1.68 A resolution range. The final model contained 1165 protein atoms for all of the 148 residues, four Ca(2+) ions, and 172 water molecules. The dumbbell structure has seven alpha-helices including a long 7.8 turn central helix connecting the two terminal domains each containing two EF-hand (helix-loop-helix motif) calcium-binding sites. The loops within each pair of EF-hand motifs in the N- and C-terminal domains are brought into juxtaposition to form a pair of hydrogen-bonded antiparallel beta-sheets which are extended at either ends by water bridges. The four calcium-binding EF-hands are superposable with r.m.s. deviations of 0.31-0.79 A. The best agreement is between site 1 and site 3 and the worst agreement is between site 1 and 4. The largest differences are in the ninth and tenth residues of the calcium-binding loops probably because of their involvement in the mini beta-sheets. The calcium coordination distances vary between 2.04 and 2.69 A, average 2.34 A. The rPCaM and wild-type PCaM have an r.m.s. deviation of 0.36 A for equivalent C(alpha) atoms. The side chains of Lys13 and Lys115 are more extended in rPCaM compared to the wild type where the post-translational modified di- and tri-methylated lysine residues are more folded. The sequence of PCaM differs from those of mammalian (MCaM) and Drosophila calmodulin (DCaM), but the overall structures are very similar, with r.m.s,. deviations of 0.44 and 1.68 A for equivalent C(alpha) atoms, respectively. However, in rPCaM, the first four N-terminal residues stretch out and make intermolecular crystal contacts, in contrast to those in recombinant Drosophila calmodulin (rDCaM), they stretch out in the opposite direction and towards the second calcium-binding site (see note below), while in MCaM and wild-type PCaM, the N-terminal residues are not visible. The central helix in rPCaM has all its backbone hydrogen bonds intact with no unusually long separation between the carbonyl and amide groups as found in MCaM and rDCaM.
- Laboratory of Biological Macromolecular Structure, Department of Chemistry, Biochemistry and Biotechnology Center, The Ohio State University, Columbus, OH 43210, USA.
Organizational Affiliation: