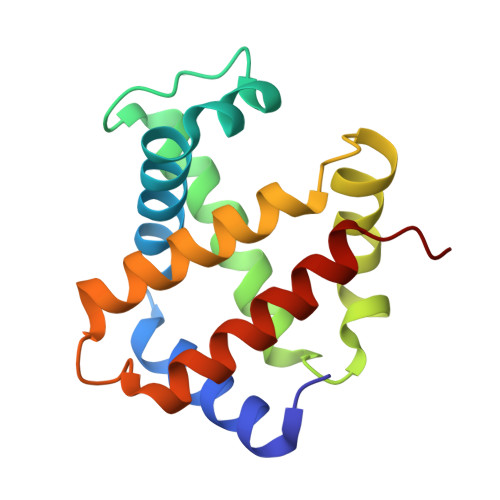
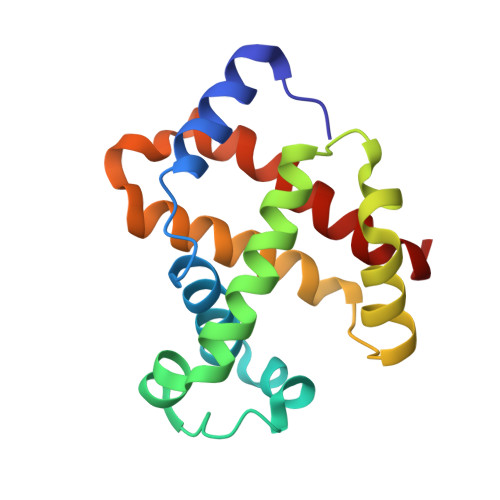
Crystal structures of HbA2 and HbE and modeling of hemoglobin delta4: interpretation of the thermal stability and the antisickling effect of HbA2 and identification of the ferrocyanide binding site in Hb.
Sen, U., Dasgupta, J., Choudhury, D., Datta, P., Chakrabarti, A., Chakrabarty, S.B., Chakrabarty, A., Dattagupta, J.K.(2004) Biochemistry 43: 12477-12488
- PubMed: 15449937 Search on PubMed
- DOI: https://doi.org/10.1021/bi048903i
- Primary Citation Related Structures:
1SHR, 1SI4 - PubMed Abstract:
Hemoglobin A(2) (alpha(2)delta(2)) is an important hemoglobin variant which is a minor component (2-3%) in the circulating red blood cells, and its elevated concentration in beta-thalassemia is a useful clinical diagnostic. In beta-thalassemia major, where there is beta-chain production failure, HbA(2) acts as the predominant oxygen deliverer. HbA(2) has two more important features. (1) It is more resistant to thermal denaturation than HbA, and (2) it inhibits the polymerization of deoxy sickle hemoglobin (HbS). Hemoglobin E (E26K(beta)), formed as a result of the splice site mutation on exon 1 of the beta-globin gene, is another important hemoglobin variant which is known to be unstable at high temperatures. Both heterozygous HbE (HbAE) and homozygous HbE (HbEE) are benign disorders, but when HbE combines with beta-thalassemia, it causes E/beta-thalassemia which has severe clinical consequences. In this paper, we present the crystal structures of HbA(2) and HbE at 2.20 and 1.74 A resolution, respectively, in their R2 states, which have been used here to provide the probable explanations of the thermal stability and instability of HbA(2) and HbE. Using the coordinates of R2 state HbA(2), we modeled the structure of T state HbA(2) which allowed us to address the structural basis of the antisickling property of HbA(2). Using the coordinates of the delta-chain of HbA(2) (R2 state), we also modeled the structure of hemoglobin homotetramer delta(4) that occurs in the case of rare HbH disease. From the differences in intersubunit contacts among beta(4), gamma(4), and delta(4), we formed a hypothesis regarding the possible tetramerization pathway of delta(4). The crystal structure of a ferrocyanide-bound HbA(2) at 1.88 A resolution is also presented here, which throws light on the location and the mode of binding of ferrocyanide anion with hemoglobin, predominantly using the residues involved in DPG binding. The pH dependence of ferrocyanide binding with hemoglobin has also been investigated.
- Crystallography and Molecular Biology Division, Saha Institute of Nuclear Physics, 1/AF Bidhan Nagar, Kolkata 700064, India.
Organizational Affiliation: