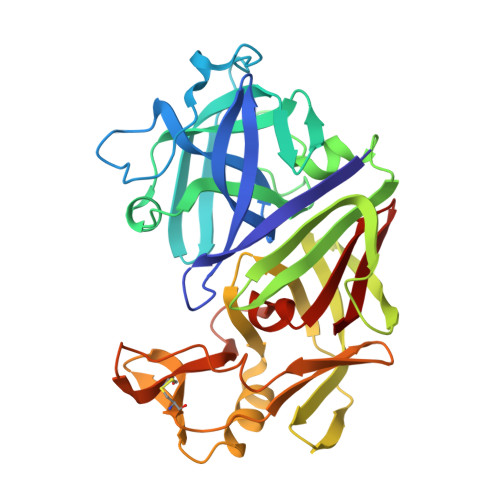
X-ray analyses of aspartic proteinases. The three-dimensional structure at 2.1 A resolution of endothiapepsin.
Blundell, T.L., Jenkins, J.A., Sewell, B.T., Pearl, L.H., Cooper, J.B., Tickle, I.J., Veerapandian, B., Wood, S.P.(1990) J Mol Biology 211: 919-941
- PubMed: 2179568 Search on PubMed
- DOI: https://doi.org/10.1016/0022-2836(90)90084-Y
- Primary Citation Related Structures:
1ENT - PubMed Abstract:
The molecular structure of endothiapepsin (EC 3.4.23.6), the aspartic proteinase from Endothia parasitica, has been refined to a crystallographic R-factor of 0.178 at 2.1 A resolution. The positions of 2389 protein non-hydrogen atoms have been determined and the present model contains 333 solvent molecules. The structure is bilobal, consisting of two predominantly beta-sheet domains that are related by an approximate 2-fold axis. Of approximately 170 residues, 65 are topologically equivalent when one lobe is superimposed on the other. Twenty beta-strands are arranged as five beta-sheets and are connected by regions involving 29 turns and four helices. A central sheet involves three antiparallel strands from each lobe organized around the dyad axis. Each lobe contains a further local dyad that passes through two sheets arranged as a sandwich and relates two equivalent motifs of four antiparallel strands (a, b, c, d) followed by a helix or an irregular helical region. Sheets 1N and 1C, each contain two interpenetrating psi structures contributed by strands c,d,d' and c',d',d, which are related by the intralobe dyad. A further sheet, 2N or 2C, is formed from two extended beta-hairpins from strands b,c and b',c' that fold above the sheets 1N and 1C, respectively, and are hydrogen-bonded around the local intralobe dyad. Asp32 and Asp215 are related by the interlobe dyad and form an intricate hydrogen-bonded network with the neighbouring residues and comprise the most symmetrical part of the structure. The side-chains of the active site aspartate residues are held coplanar and the nearby main chain makes a "fireman's grip" hydrogen-bonding network. Residues 74 to 83 from strands a'N and b'N in the N-terminal lobe form a beta-hairpin loop with high thermal parameters. This "flap" projects over the active site cleft and shields the active site from the solvent region. Shells of water molecules are found on the surface of the protein molecule and large solvent channels are observed within the crystal. There are only three regions of intermolecular contacts and the crystal packing is stabilized by many solvent molecules forming a network of hydrogen bonds. The three-dimensional structure of endothiapepsin is found to be similar to two other fungal aspartic proteinases, penicillopepsin and rhizopuspepsin. Even though sequence identities of endothiapepsin with rhizopuspepsin and penicillopepsin are only 41% and 51%, respectively, a superposition of the three-dimensional structures of these three enzymes shows that 237 residues (72%) are within a root-mean-square distance of 1.0 A.
- Department of Crystallography, Birkbeck College, University of London, England.
Organizational Affiliation: