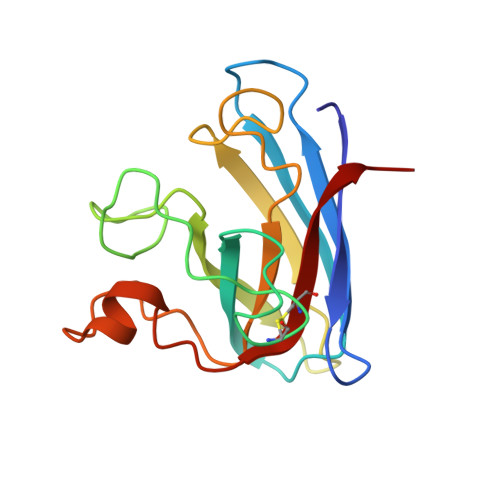
Solution structure of reduced monomeric Q133M2 copper, zinc superoxide dismutase (SOD). Why is SOD a dimeric enzyme?.
Banci, L., Benedetto, M., Bertini, I., Del Conte, R., Piccioli, M., Viezzoli, M.S.(1998) Biochemistry 37: 11780-11791
- PubMed: 9718300 Search on PubMed
- DOI: https://doi.org/10.1021/bi9803473
- Primary Citation Related Structures:
1BA9 - PubMed Abstract:
Copper, zinc superoxide dismutase is a dimeric enzyme, and it has been shown that no cooperativity between the two subunits of the dimer is operative. The substitution of two hydrophobic residues, Phe 50 and Gly 51, with two Glu's at the interface region has disrupted the quaternary structure of the protein, thus producing a soluble monomeric form. However, this monomeric form was found to have an activity lower than that of the native dimeric species (10%). To answer the fundamental question of the role of the quaternary structure in the catalytic process of superoxide dismutase, we have determined the solution structure of the reduced monomeric mutant through NMR spectroscopy. Another fundamental issue with respect to the enzymatic mechanism is the coordination of reduced copper, which is the active center. The three-dimensional solution structure of this 153-residue monomeric form of SOD (16 kDa) has been determined using distance and dihedral angle constraints obtained from 13C, 15N triple-resonance NMR experiments. The solution structure is represented by a family of 36 structures, with a backbone rmsd of 0.81 +/- 0.13 A over residues 3-150 and of 0.56 +/- 0.08 A over residues 3-49 and 70-150. This structure has been compared with the available X-ray structures of reduced SODs as well as with the oxidized form of human and bovine isoenzymes. The structure contains the classical eight-stranded Greek key beta-barrel. In general, the backbone and the metal sites are not affected much by the monomerization, except in the region involved in the subunit-subunit interface in the dimeric protein, where a large disorder is present. Significative changes are observed in the conformation of the electrostatic loop, which forms one side of the active site channel and which is fundamental in determining the optimal electrostatic potential for driving the superoxide anions to the copper site which is the rate-limiting step of the enymatic reaction under nonsaturating conditions. In the present monomer, its conformation is less favorable for the diffusion of the substrate to the reaction site. The structure of the copper center is well-defined; copper(I) is coordinated to three histidines, at variance with copper(II) which is bound to four histidines. The hydrogen atom which binds the histidine nitrogen detached from copper(I) is structurally identified.
- Department of Chemistry, University of Florence, Italy.
Organizational Affiliation: