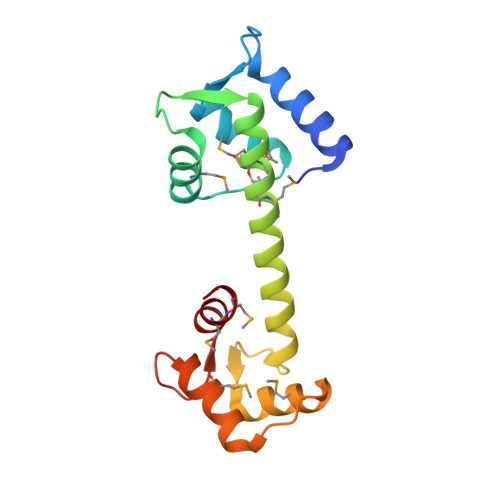
Structure of a Trapped Intermediate of Calmodulin: Calcium Regulation of EF-hand Proteins from a New Perspective.
Grabarek, Z.(2005) J Mol Biol 346: 1351-1366
- PubMed: 15713486
- DOI: https://doi.org/10.1016/j.jmb.2005.01.004
- Primary Citation of Related Structures:
1Y6W - PubMed Abstract:
Calmodulin (CaM) is a multifunctional Ca2+-binding protein that regulates the activity of many enzymes in response to changes in the intracellular Ca2+ concentration. There are two globular domains in CaM, each containing a pair of helix-loop-helix Ca2+-binding motifs called EF-hands. Ca2+-binding induces the opening of both domains thereby exposing hydrophobic pockets that provide binding sites for the target enzymes. Here, I present a 2.4 A resolution structure of a calmodulin mutant (CaM41/75) in which the N-terminal domain is locked in the closed conformation by a disulfide bond. CaM41/75 crystallized in a tetragonal lattice with the Ca2+ bound in all four EF-hands. In the closed N-terminal domain Ca ions are coordinated by the four protein ligands in positions 1, 3, 5 and 7 of the loop, and by two solvent ligands. The glutamate side-chain in the 12th position of the loop (Glu31 in site I and Glu67 in site II), which in the wild-type protein provides a bidentate Ca2+ ligand, remains in a distal position. Based on a comparison of CaM41/75 with other CaM and troponin C structures a detailed two-step mechanism of the Ca2+-binding process is proposed. Initially, the Ca2+ binds to the N-terminal part of the loop, thus generating a rigid link between the incoming helix (helix A, or helix C) and the central beta structure involving the residues in the sixth, seventh and eighth position of the loop. Then, the exiting helix (helix B or helix D) rotates causing the glutamate ligand in the 12th position to move into the vicinity of the immobilized Ca2+. An adjustment of the phi, psi backbone dihedral angles of the Ile residue in the eighth position is necessary and sufficient for the helix rotation and functions as a hinge. The model allows for a significant independence of the Ca2+-binding sites in a two-EF-hand domain.
Organizational Affiliation:
Boston Biomedical Research Institute, Watertown, MA 02472, USA. grabarek@bbri.org